 联系我们
联系我们因此重组抗体表达这项技术的研发也是基于抗体研究领域的科学家对于抗体多样性研究和理解的基础上,而普健生物也经过多年的抗体表达实践和对哺乳系统表达的优化,研发出一项高效表达重组抗体的专有技术。可经过本文来了解一下重组抗体表达技术以及抗体结构和抗体多样性产生的机制。
重组抗体表达
普健生物自有的哺乳表达系统高产量pATX系列载体及驯化293系列和CHO系列细胞,能实现全长抗体以及多种形式的重组抗体片段的高效分泌表达;无需免疫和筛选,直接利用抗体基因序列即可实现重组抗体的高产和可重复生产。
诸多表达优势
1. 抗体药物研发专家和技术团队,提供个性化抗体发现整体方案,重组抗体表达量高,采用高密度转染,可实现瞬转表达量几百毫克每升,稳转可优化到克级。
2. 抗体表达经验丰富,已成功表达过多种形式的全长抗体(Human IgG, mouse IgG, human Ig A, IgM, IgE, mouse IgM,rabbit IgG 等)和抗体片段(scFv,Fab,tandem-scfv,VHH,Fc融合蛋白,bi-specific antibody等)。
3. 一站式服务:拥有多种服务平台,在实现重组抗体设计表达和纯化的同时,进行抗体质控和活性验证,并由经验丰富的专家根据客户需求进行人源化设计验证。
4. 自主知识产权的GS敲除系统细胞系,长期稳定表达,抗体质量更稳定。
5. 周期短,可提供2~3周急速交付重组抗体表达服务。
抗体的结构及多样化过程
典型的抗体分子由重链和轻链组成,形成的免疫球蛋白结构域又可以分为可变区和恒定区,分别负责抗原的结合和下游免疫反应的激活。
在B细胞发育早期阶段,编码可变区的V元件、J元件,以及重链可变区特有的D元件,可以通过核酸内切酶RAG起始的V(D)J重排进行组合和连接从而形成可变区基因外显子,所分泌的低亲和力IgM抗体构成了初始的抗体库。在外周淋巴组织结构生发中心中,B细胞特异性DNA去氨酶AID能够起始可变区高频突变和重链恒定区抗体类型转换,从而形成了表达高亲和力不同抗体类型的抗体库。

典型的抗体分子和抗体多样化过程
体类型转换的分子机制
抗体类型转换是抗体多样化过程中的重要一步,其产生的不同类型抗体能够激活不同的下游免疫反应。
B细胞特异性DNA去氨酶AID起始了这一过程,产生的DNA断裂则由细胞内DNA损伤修复系统进一步连接完成整个过程。遗传学证据清晰地表明,抗体类型转换由两步组成:第一步,AID介导的双链DNA断裂的产生;第二步,双链DNA断裂激活DNA损伤应答机制将两个双链DNA断裂连接起来。
1. AID靶向基因组DNA底物引起双链DNA断裂的机制

A:AID去氨基酶活性示意图。AID作用于N4位点,能够将单链DNA上的胞苷(C)残基脱氨产生尿苷(U);B、C、D:基因组转录过程中产生的转录泡、负超螺旋结构以及R-环可以提供单链DNA招募AID;深色线条和浅色线条分别代表DNA双链的模板链和非模板链,红线代表RNA转录产物。E:AID相互作用因子RPA复合物和RNA外切体复合物能够为AID提供单链DNA底物。F:对向转录招募AID到非免疫球蛋白基因靶点:蓝线代表转录, 红线代表反转录。
2. 双链DNA损伤修复与S区断裂末端连接
抗体类型转换中产生的S区的双链DNA断裂的修复,并不依赖于AID去氨酶活性。AID起始形成S区的双链DNA断裂可以激活细胞内通用的双链DNA损伤应答系统,最终非同源DNA末端连接系统将两个S区断裂连接起来。
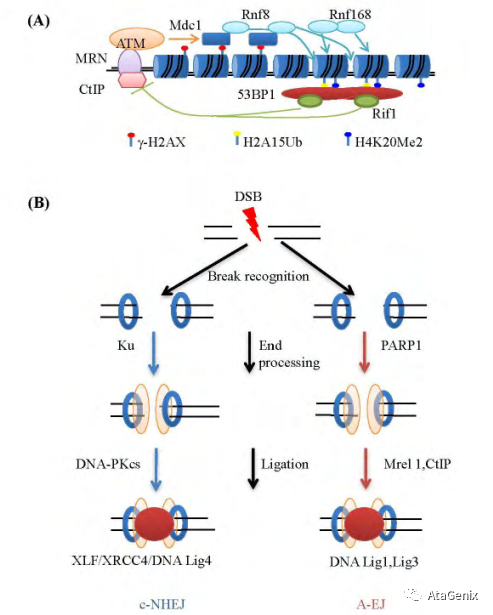
DNA末端修复连接途径
A: ATM依赖的双链DNA损伤应答机制。MRN复合体可以识别DSB末端招募ATM并激活ATM激酶活性, 使多种下游蛋白包括H2AX、53BP1等发生磷酸化。磷酸化的H2AX即γ-H2AX可以进一步招募其他双链DNA损伤应答因子MDC1, RNF8能够与磷酸化的MDC1相互作用, 并泛素化修饰组蛋白H1。RNF168可以识别组蛋白H1泛素化修饰, 并进一步泛素化修饰组蛋白H2A。53BP1识别组蛋白H2A泛素化修饰而进一步招募至DNA损伤位点。ATM磷酸化的53BP1能够招募Rif1进一步抑制由CtIP介导的双链DNA末端的单链切除, 从而使DNA断裂末端更容易被非同源DNA末端连接途径修复并同时抑制同源重组的发生。B:在c-NHEJ过程中,Ku结合至DSB末端, DNA-PKcs被进一步招募至DNA的DSB上,XRCC4/Lig4复合物最终连接两个末端。A-EJ途径中, PARP1可能参与了对DSB的识别, Mrel 1和CtIP可能介导末端连接, 最终由Lig1或Lig3进行末端连接。
重组抗体表达的相关研究进展
抗体产生和多样化过程的缺陷在临床上表现为各种免疫缺陷疾病,而抗体多样化的失控则可能造成B细胞淋巴瘤的产生。
抗体多样化中,抗体基因在基因组水平面临DNA修饰和损伤的发生。这种DNA水平的损伤虽然受到精细地调控,但往往也造成了基因组的不稳定性导致癌症的发生。生发中心B细胞来源的B细胞淋巴瘤中,AID起始的DNA损伤产生的染色体易位、基因突变等是癌变的主要机制之一。研究AID靶向机制和抗体多样化中DNA损伤修复机制,可以为生发中心来源的B细胞淋巴瘤,如滤泡型淋巴瘤、弥漫大B细胞淋巴瘤等,以及CLL的治疗和诊断提供新的思路和技术手段。
目前我们对抗体多样化机制的认识仍存在大量的空白,致力于抗体类型的研究从未停歇。通过基因工程对编码抗体的基因按不同需要进行加工改造和重新装配,转染适当的受体细胞从而表达抗体,这种通过基因工程重组表达抗体的技术,现已广泛应用于抗体药物研究。
重组抗体的优势
重组抗体包括嵌合抗体、人源化抗体、小分子抗体、双特异性抗体等。与传统的单克隆抗体和多克隆抗体相比,重组抗体具有独特的优势,现已被广泛地应用在科学研究、免疫诊断及治疗性抗体领域。在被广泛应用的情况下,重组抗体表达技术水平也在日益提高,重组抗体除了以上优势之外还有以下一些优点。
(1)重组抗体具有序列已知、抗体基因可以长期保存、抗体性能稳定、实验重复性好等优点,而单克隆细胞株可能存在抗体染色体丢失、细胞复苏后停止生长或死亡等风险。
(2)可以进行基因工程改造:重组抗体可以进行重组改造,包括抗体片段表达,同型置换,嵌合抗体,抗体人源化等。
(3)可以大量制备抗体:重组抗体可以快速实现抗体的大量制备,不需要使用动物,克服使用动物的伦理问题。









