 联系我们
联系我们在生命科学领域中,有两种极为重要的生物大分子一直在被科学家们研究,一种是执行大部分生命活动的蛋白质,一种是承载遗传信息的核酸。在噬菌体展示技术出现以前,无论是成本、通量抑或效率,解码核酸序列的能力远优于解码蛋白质序列的能力。科学家们利用噬菌体展示技术将蛋白质相关信息转化为核酸信息后,通过高通量测序来实现对蛋白质信息的解读,这也为蛋白质研究提供了更为便捷的方法。

噬菌体展示技术其原理在于将展示在其表面上的蛋白质/抗体或者小肽的序列与编码它们的核酸序列进行关联,利用噬菌体的超强扩增能力构建小肽或者抗体文库,多样性可以达到109以上;然后利用噬菌体展示技术通过多轮淘选得到针对特定对象的亲和肽或者抗体。
那么噬菌体展示技术是如何发现和发展的呢?这首先得从噬菌体开始说起。
噬菌体是一类病毒,顾名思义是吞噬细菌、真菌、超级细菌等微生物的病毒。自然界中,噬菌体无处不在,以其极端的多样性持续颠覆生命领域的“游戏规则”,被生物科学家成为生命宇宙的“暗物质”,也被开发成为病原微生物防控的“核武器”。除此之外,噬菌体展示是其应用的一个重要领域。
1985年,G. P. Smith教授将EcoR I基因片段插入丝状噬菌体f1的BamH I位点,转化E. coli,通过与f1噬菌体的结构蛋白融合,成功将EcoR I(“颜值”)展示于噬菌体的表面(1)。此后,随着大量噬菌体展示文库的构建、遗传操作体系的建立以及展示平台与体系的拓展,噬菌体展示技术在不同领域,尤其是蛋白质和抗体相关领域,发挥了重要作用。
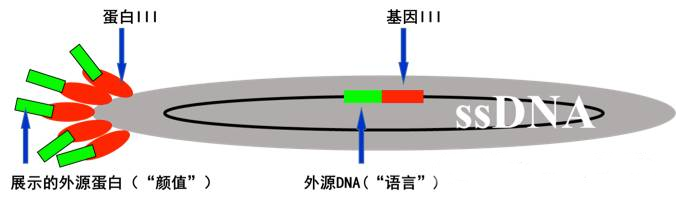
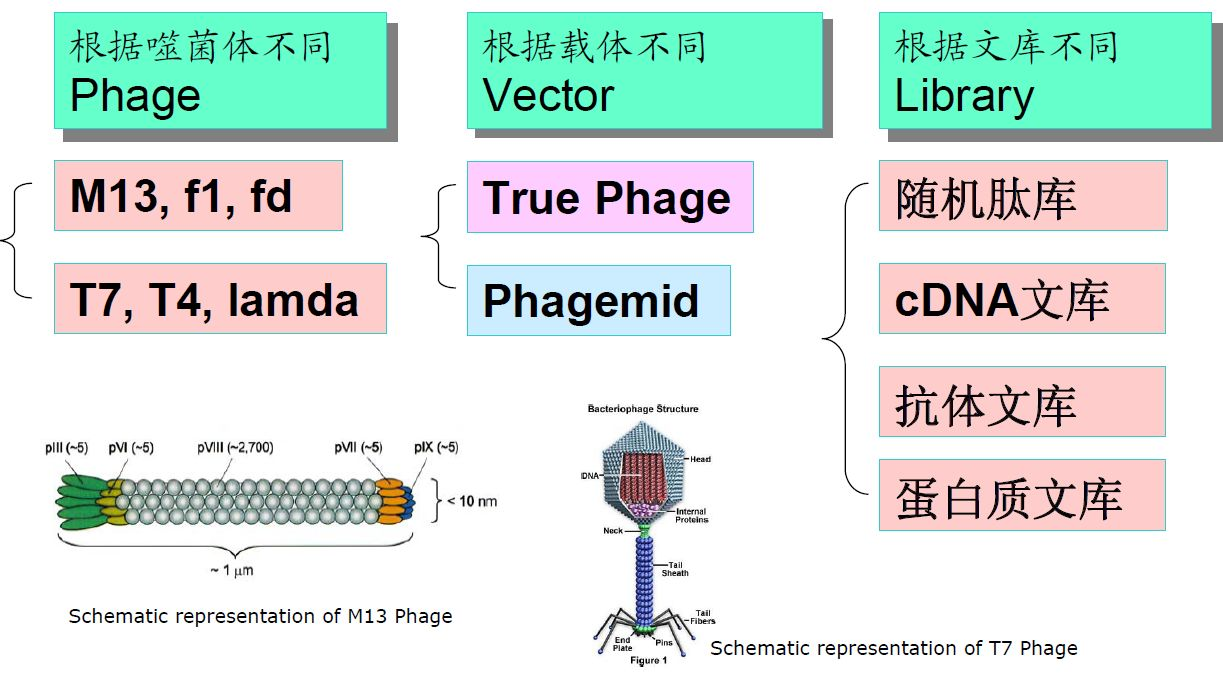
噬菌体展示技术有多种分类标准。根据噬菌体的不同,可以分为M13、T7、T4、λ等多种体系;根据载体的不同,可以分为噬菌体载体(True Phage)和噬菌粒载体(Phagemid);根据文库的不同,可以分为随机肽库、cDNA文库、抗体文库、蛋白质文库等。
M13噬菌体展示系统
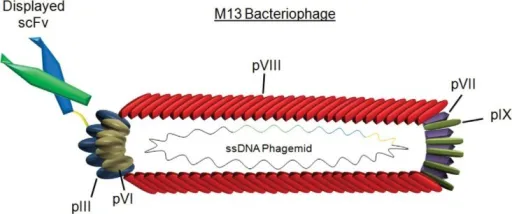
单链丝状噬菌体(M13)属于丝状噬菌体,含有单链DNA基因组,其衣壳蛋白为管状,通常由几千个拷贝的主要衣壳蛋白(PⅧ)和位于尾端的次要衣壳蛋白(PⅢ)组成。由于该蛋白的分子量很小,只适合用来展示外源短肽。外源肽段的太大会影响病毒包装,不能形成有功能的噬菌体。该系统比较适合用来筛选低亲和力的配体。
T7噬菌体展示系统
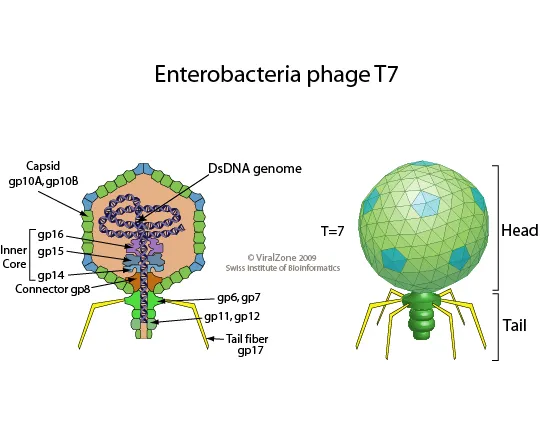
T7 噬菌体基因组为线性双链DNA,其衣壳蛋白通常有两种形式,即10A(344个氨基酸残基)和10B(397个氨基酸残基),10B衣壳蛋白区存在于噬菌体表面,所以被用来构建噬菌体展示系统。与M13系统相比,T7噬菌体直接使宿主菌进行裂解,不需经过分泌过程,广泛应用于筛选不同分子量,不同亲和力的蛋白质。
T4噬菌体展示系统
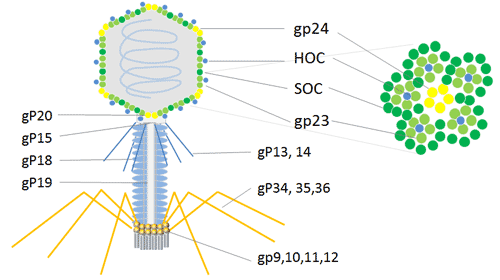
T4噬菌体基因组DNA为双链线形,呈环状排列,噬菌体衣壳的有两种非必需外壳蛋白SOC(small outer capsid protein,9 ku)和HOC(highly antigenic outer capsid protein,40 ku),因此它表达的蛋白不需要复杂的蛋白纯化,可展示各种大小的多肽或蛋白质,很少受到限制。
λ噬菌体展示系统
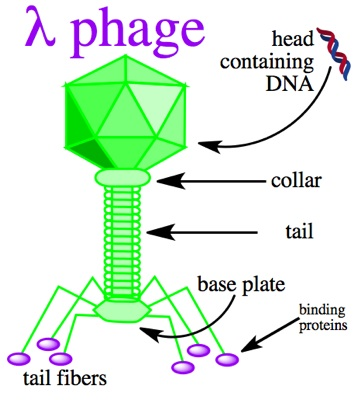
λ噬菌体是长尾噬菌体科的一种温和噬菌体,有直径55nm的二十面体头部,末端有细长尾丝。基因组为48.5 kb的线性双链DNA分子,有黏性末端即单链延伸12个核苷酸,感染后线性基因组可立即环化。噬菌体的头部由D蛋白和V蛋白构成,可以构建D蛋白和V蛋白的展示系统。λ噬菌体是在宿主细胞内完成装配的,无需将外源肽或蛋白分泌到细菌胞膜外,可展示有活性的大分子蛋白质(100 kDa以上)及宿主细胞有毒性的蛋白质,适用范围极广。









