 联系我们
联系我们展示技术,尤其是噬菌体展示技术,在许多领域得到了广泛的应用。展示技术的理论核心是噬菌体表面的蛋白质/肽与同一噬菌体内部的编码DNA序列之间的物理联系。从噬菌体展示的肽/蛋白/抗体库出发,利用下一代测序技术(NGS)日益强大的DNA测序/解码能力,可以很容易地以高通量的方式获得丰富的蛋白质相关信息。基于这些信息,许多科学和临床问题可以很容易地解决。在过去的几年里,在NGS、液滴技术和大规模寡核苷酸合成的帮助下,我们已经见证了噬菌体展示技术在技术开发和应用方面的巨大进步。今天小编将分享当前噬菌体展示技术的最新进展。
1985年,Smith将EcoRI基因片段插入噬菌体f1的基因组中得到了最终的噬菌体,命名为fECO1,它保留了噬菌体的传染性和外源蛋白EcoRI对抗体的可及性。fECO1的诞生标志着一种新的技术噬菌体展示技术被发明,并且正是由于这一进展,Smith教授获得2018年诺贝尔化学奖。
作为展示技术的核心,感兴趣的蛋白质在物理上与其编码的DNA片段相连。与解码蛋白质序列的方法相比,DNA测序技术要先进得多。利用下一代测序(NGS)的强大功能,可以通过NGS的展示平台回答蛋白质相关的问题。除了Smith所描述的噬菌体展示系统,更多其他的展示平台也逐渐被开发出来,如基于Ff噬菌体、λ噬菌体、T7噬菌体、T4噬菌体、大肠杆菌、植物乳杆菌、酵母等展示系统;mRNA展示技术;核糖体展示系统;和SNAP展示系统等。迄今为止,已经发表了许多关于展示平台的综述文章和书籍,涵盖了展示平台的各个方面,如基于Ff噬菌体的展示系统,基于Ff噬菌体的载体,以及展示库建设的策略。接下来,我们将主要关注噬菌体展示技术的最新进展和新应用。
生物学的噬菌体展示
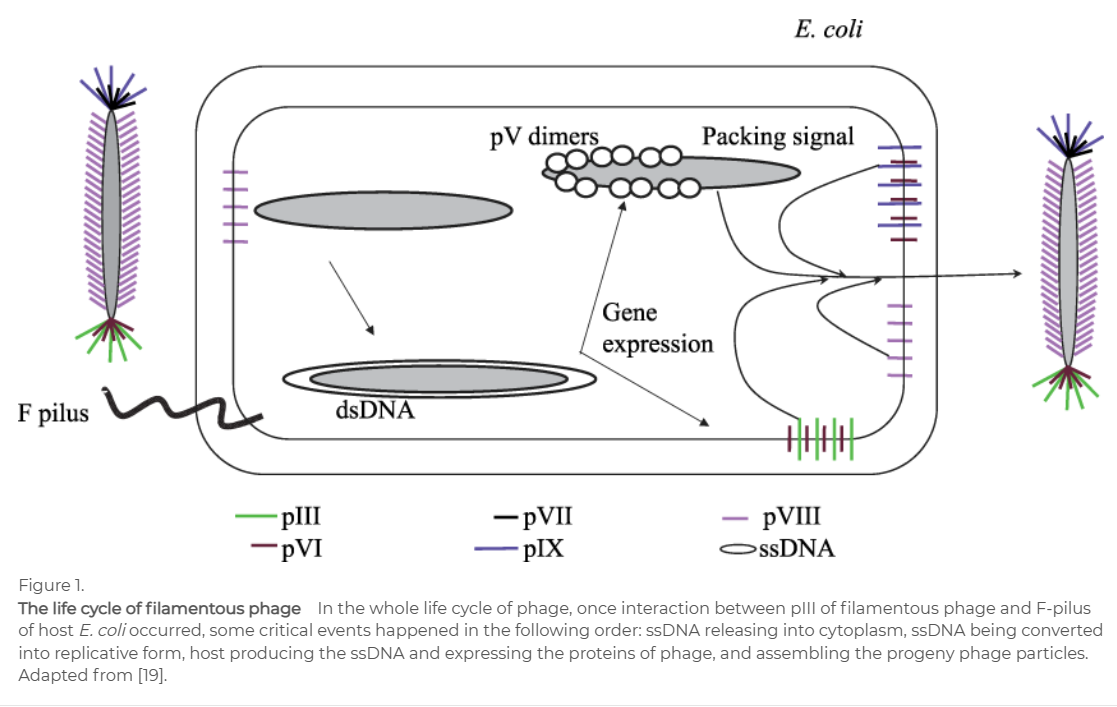
为了更清楚地介绍噬菌体展示技术和应用的最新进展,首先我们需要了解噬菌体的生命周期。以被广泛应用丝状噬菌体为例,如下图所示,丝状噬菌体由五个结构蛋白(pIII, pVI, pVII, pVIII, pIX)和一个ssDNA组成。在野生型丝状噬菌体中,一个末端有5个pIII和pVI,另一个末端有5个pVII和pIX。剩下的pVIII,每个噬菌体约2700拷贝,与ssDNA相互作用。基因组长度为6407 bp。在丝状噬菌体生命周期的早期,噬菌体的pIII与大肠杆菌的f -菌毛相互作用。一旦相互作用发生,f -菌毛开始解聚,噬菌体被拉入宿主。随后,ssDNA被释放到宿主中。将双链DNA转化为复制形式后,作为模板生成ssDNA并表达用于组装子代噬菌体颗粒的蛋白质。在噬菌体展示中,外源蛋白的基因片段插入到基因组中,与结构蛋白的基因融合。结果,外源蛋白与相应的结构蛋白融合。融合蛋白和ssDNA被组装成相同的子代噬菌体粒子。由于基因与外源蛋白质之间存在物理联系,可以通过基因序列间接获得蛋白质的序列。理论上,所有的结构蛋白都可以作为与外源蛋白融合的载体蛋白。而丝状噬菌体的pIII和pVIII通常被用作载体蛋白。
噬菌体显示技术的最新进展
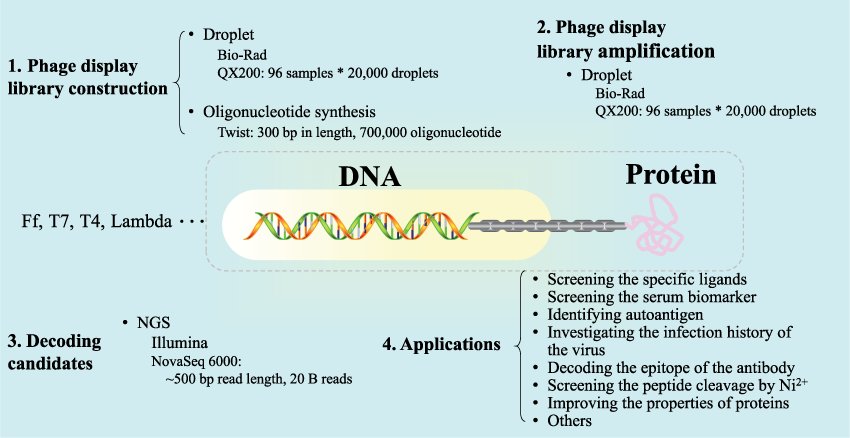
噬菌体展示技术因其独特的特性被广泛应用于筛选配体、鉴定表位、改善目标分子性能等领域。近年来,如图所示,随着NGS、液滴、大量寡核苷酸合成等相关技术的发展,噬菌体展示技术得到了加速发展。
噬菌体展示的最新应用
通过噬菌体展示,可以将显示库中符合定义标准的克隆,如具有较高的亲和力或较高的酶活性。近年来,噬菌体展示除了应用于筛选目标分子或组织的特异性配体外,还被进一步应用于血清生物标志物的筛选、抗原的鉴定、病毒感染史的调查、抗体表位的解码、Ni2+对肽切割的筛选、蛋白性质的改善等领域。
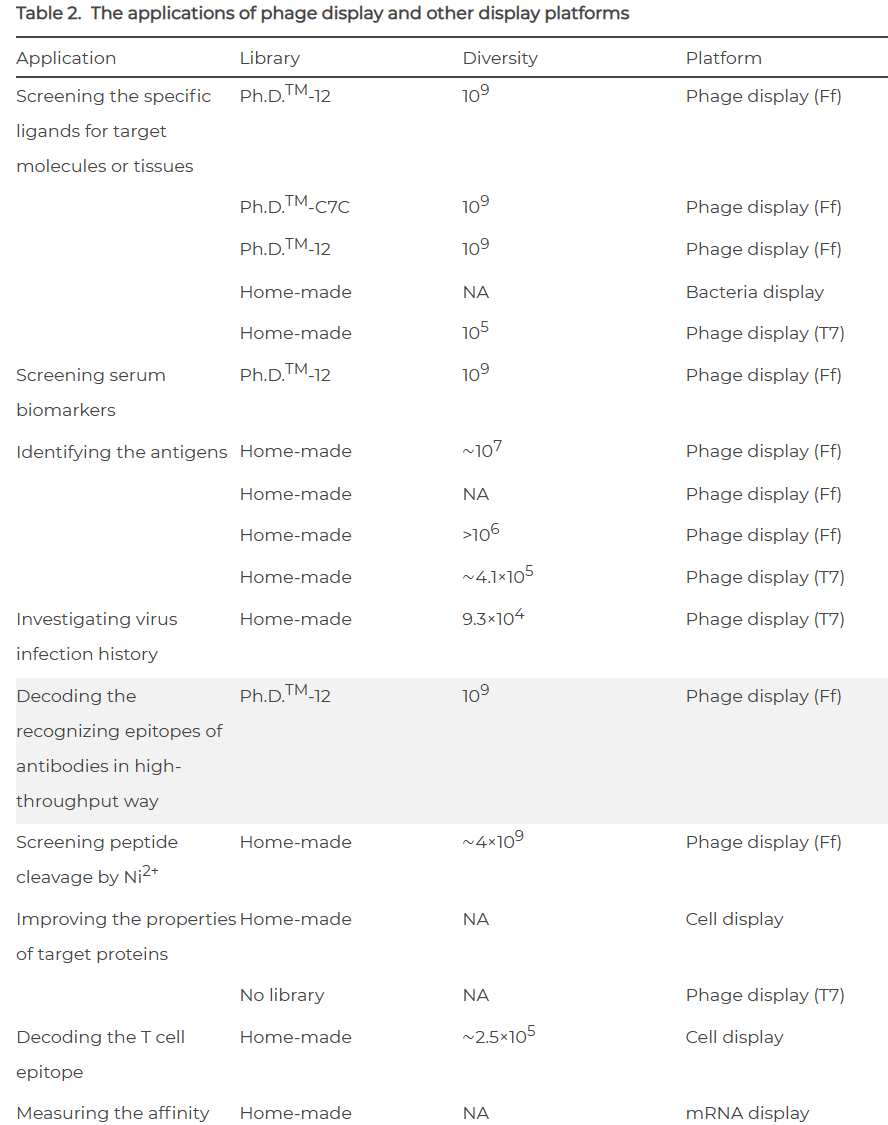
考虑到DNA和蛋白质解码的不同,噬菌体展示是通过解码相应DNA获得蛋白质氨基酸序列信息的理想平台。近年来,随着NGS技术、液滴技术和大量寡核苷酸合成技术的发展,噬菌体展示技术在构建噬菌体文库、解码显示元素、评估文库质量等方面取得了很大进展。然而,噬菌体展示领域仍然存在一些挑战。毫无疑问,噬菌体展示将在不久的将来成为连接蛋白质和相应DNA的更理想的平台。
转载说明:本文图文源自ACTA BIOCHIMICA ET BIOPHYSICA SINICA,仅做学术分享使用;感谢每一位作者的辛苦付出与创作,除转载众多无法溯源的文章,我们均在文章中备注了来源。如转载涉及版权等问题,请联系我们删除,非常感谢!









