 联系我们
联系我们自1965年发现B细胞以来(Cooper et al. 1965),我们对B细胞的发育、成熟和功能的认识有了巨大的进步。B细胞通过抗原呈递、细胞因子分泌和自身抗体的产生,是多种自身免疫性疾病发病机制的核心。
B淋巴细胞通过产生保护性抗体在对抗病原体和肿瘤的免疫反应中起着至关重要的作用。它们也与自身免疫的发生和控制有关,其中免疫反应指向自身抗原。不同的B细胞群表现出保护和自我毁灭的能力。目前了解最多的B淋巴细胞是介导适应性体液反应的传统骨髓来源的B2细胞。在包括类风湿关节炎(RA)和系统性红斑狼疮(SLE)在内的各种自身免疫性疾病中,B2细胞负责产生自身抗体、将内源性肽提呈给自身反应性T细胞以及分泌促炎细胞因子。除了B2细胞外,还有一群独特的分泌天然抗体的淋巴细胞,称为B1细胞。B1细胞产生于胎儿肝脏,在腹膜中发现,并具有静息自我更新的能力。B1细胞产生针对细菌多糖抗原的天然IgM,并表现出显著的自我反应性。B1细胞产生的自身抗体和细胞因子可介导自身免疫。然而,B1细胞来源的自身抗体也有助于清除凋亡细胞(自身抗原的主要来源),并且似乎可促进肠道稳态。同时,调节性B细胞(regulatory B cells, Bregs)通过分泌IL-10、TGF -β等免疫调节细胞因子,对自身反应性淋巴细胞发挥接触依赖性抑制作用,在耐受发生中发挥明确的作用。
自身免疫中B细胞功能障碍的原因尚不完全明确。虽然代谢是细胞活动的明确调节因素,但直到最近才被认为是健康和疾病中B细胞功能的决定因素。生理上,代谢使B细胞正常发育、分化和抗体分泌。不出所料,不同发育和成熟阶段的B细胞的不同表型反映在代谢活性的显著差异上。此外,不同的B细胞亚群,特别是B1和B2细胞,利用不同的代谢途径。代谢是抗自身免疫检查点缺失和无力的重要组成部分,代谢失调与逃避耐受性检查点和自身反应性B细胞功能增强相关。除了传统的细胞代谢外,B细胞还依赖于自噬,这是一种主要对受损细胞成分进行降解处理的机制。自噬维持营养剥夺期间的代谢稳态,并支持浆细胞(PC)的长期活力。病理学上,自噬似乎支持自身反应性B细胞破坏自身免疫检查点,被固有免疫信号激活,并将自身抗原提呈给T淋巴细胞。
传统的B2 B细胞的发育必须产生大量的具有抗体分泌能力的细胞库,同时清除具有自身反应性抗原特异性的细胞。此外,快速生长和增殖必须在代谢困难的骨髓环境中实现,需要仔细平衡合成代谢和分解代谢信号传导。前者主要由c-Myc和线粒体雷帕霉素靶蛋白复合物(mTORC)信号通路介导,通过上调糖酵解和氧化磷酸化(OXPHOS)来促进蛋白质合成和细胞生长。mTORC信号通路在B细胞发育过程中起着至关重要的作用。在小鼠中,敲除mtorc1相关蛋白Raptor可阻止白细胞介素(IL) - 7驱动的前b细胞发育。没有mTORC1信号传导,前b细胞向前b细胞转化的能力较低,而前b细胞是表达前b细胞受体(BCR)的前体,BCR由成熟的免疫球蛋白重链和替代轻链组成。与mTORC1不同,mTORC2在早期B细胞发育中的作用一直存在争议,尽管它似乎有助于外周B细胞的成熟。特别是,在B细胞发育的终末阶段,mTORC2参与调节mTORC1和c-Myc的活性。
虽然mTORC信号通路在B细胞发育过程中很重要,但过度的合成代谢活动是有害的。事实上,在Fnip1缺失后,B细胞的发育在大的前B细胞阶段受到损害。Fnip1与5 '腺苷酸活化蛋白激酶(AMPK)相互作用,AMPK是一种能量应激传感器和分解代谢调节因子,与mTORC1相互作用。虽然在Fnip1缺失的情况下,AMPK可以促进分解代谢,但其抑制mTORC信号传导的能力受损。无节制的合成代谢导致前b细胞在前bcr交联后更易发生凋亡。代谢是细胞死亡的调节因子,在抗原刺激下控制B细胞前体活力,这一概念对对抗自身免疫的耐受性检查点有意义。
大前B细胞阶段(在此期间,pre-BCR在细胞表面表达,并检测其对骨髓内自身抗原的亲和力)代表了自身免疫检查点和代谢易感性时期。尽管其在B细胞发育过程中的必要性存在争议,但葡萄糖代谢代表了这种脆弱性的一个例子。大前B细胞的快速增殖被认为是通过上调的葡萄糖代谢来维持的,因为大前B细胞比其他前体细胞需要更多的葡萄糖。大的前B细胞经历了显著的氧化应激,易受糖酵解抑制,从而损害其向小的前B细胞转化。通过自身反应性pre-BCR发出的信号可驱动磷脂酰肌醇3 -激酶(PI3K) -蛋白激酶B (Akt) -mTORC1通路的过度激活,由此产生的代谢应激可诱导负性选择。非自身反应性pre-BCR的激活不影响前B或白血病前B(pre-B ALL)细胞的Akt活化或活力。相反,自身反应性pre-BCR的激活可诱导akt依赖性快速细胞死亡。在pre-B ALL细胞中,PI3K抑制剂磷酸酶和张力蛋白同源物(PTEN)的缺失会增加糖酵解通量,但合成代谢的增加会导致ATP耗竭和细胞死亡。这些变化可被mTORC1抑制剂雷帕霉素逆转,表明自身反应性pre-BCR下游PI3K-Akt-mTORC1通路的过度激活导致能量危机。PTEN在前B细胞的发育中起着至关重要的作用,降低其对凋亡的易感性。虽然PTEN缺失影响B细胞前体的非代谢特征,如B淋巴转录因子的表达,这些结果进一步强调了平衡代谢程序在B细胞发育中的重要性。
除了控制发育的其他方面外,B淋巴细胞转录因子本身还对B细胞前体细胞施加代谢限制,这可能是为了实现过度激活诱导的细胞死亡。转录因子PAX5和IKZF1的突变在急性淋巴细胞白血病中常见,提示它们的表达可能带来选择劣势。PAX5和IKZF1在pre-B ALL细胞中的可诱导重建降低了葡萄糖摄取和ATP合成,促进了细胞死亡。值得注意的是,无功能IKZF1的B细胞特异性表达使小鼠易发生自身免疫,这支持了这些转录因子可能在B细胞发育过程中发挥耐受作用的观点。代谢受限也是B细胞无能的一个突出特征。除了凋亡和受体编辑外,耐受发生还可通过诱导无反应性作用于自身反应性B细胞及其前体细胞,使B细胞对抗原刺激反应低下。无能是早期过渡性B细胞从骨髓流出后耐受的主要机制。无反应性B细胞的特征是对BCR或toll样受体(TLR) 4刺激产生的PI3K信号传导抑制和代谢重编程受损。据推测,代谢抑制可提高无反应性自身反应性B细胞的活化阈值。
鉴于自噬在促进代谢稳态中的作用,科学家们探索了自噬在B2细胞发育中的作用。在Rag1 - / -小鼠的胎肝中,使用缺乏关键自噬基因Atg5的细胞进行重建,表明前b细胞阶段的发育受阻。当Atg5缺失局限于成熟B细胞时,脾脏和淋巴结B细胞群不受影响,这意味着自噬是B2细胞发育所必需的,而不是外周维持所必需的。然而,使用Cd79a-cre进行条理性敲除后,在Atg5表达缺失的情况下发生前b细胞向前b细胞的转化。相反,自噬是维持外周成熟B细胞群所必需的。考虑到自噬抑制剂mTORC1在发育中的关键作用,自噬在B2细胞前体细胞中可能是可控性的这一点可能并不令人惊讶。
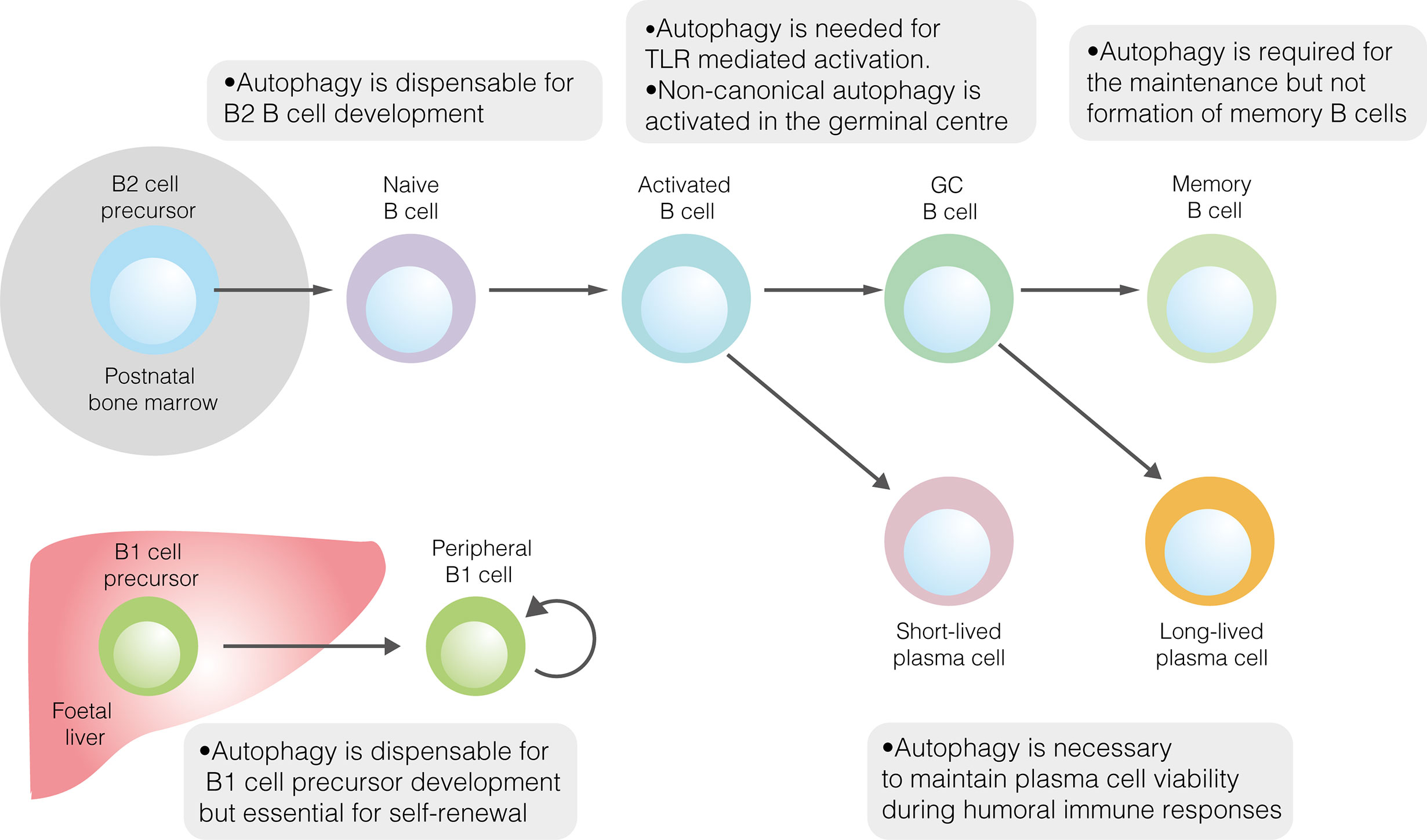
与B2细胞在骨髓中的发育相反,先天样B1细胞在胎儿肝脏中发育,然后主要迁移到腹膜和胸膜。考虑到B1细胞占据不同的生态位并依靠自我更新来维持群体,它们的代谢表型与传统B2细胞的代谢表型毫不奇怪。与滤泡B2细胞相比,腹膜B1细胞的特征是更大的葡萄糖摄取,更高的糖酵解率和OXPHOS,以及对糖酵解抑制的更高敏感性。从机制上讲,葡萄糖代谢的升高可能是由高水平的c-Myc表达驱动的。与它们在腹膜富含脂质环境中的定位一致,B1a细胞广泛获取外源性脂肪酸,同时也利用内源性脂肪酸合成。
自噬也在B1细胞中进行了研究(图1)。成熟B细胞中自噬通量的丧失会导致B1a细胞的显著消耗,但不会导致B1b细胞的显著消耗。自噬在B1细胞发育中的具体作用直到最近才被探索。在小鼠中,B1祖细胞群不受自噬缺失的影响,这表明自噬不是B1细胞发育所必需的。在没有自噬的情况下,小鼠B1a细胞群的时间变化支持了这一结论。虽然B1a细胞区室在两周龄时是正常的,但到12周龄时,它比野生型小鼠显着变小。总之,这些结果表明,自噬是外周自我更新所必需的,而不是B1a细胞的分化所必需的。在腹膜B1a细胞中,自噬似乎对控制代谢基因的表达、脂肪酸的摄取和脂滴的降解(脂噬)很重要。
参考文献
Iwan G. A. Raza,B Cell Metabolism and Autophagy in Autoimmunity,Front. Immunol., 07 June 2021,Sec. Autoimmune and Autoinflammatory Disorders,Volume 12 - 2021









